 It's tempting to oversimplify things. Like neurons. It would be nice if there were one type of neuron, and all you needed to know about how neurons work could be clearly labelled on a diagram of that one type of neuron. Well, nature LOVES to specialize. So, before getting deeper into how neurons work, I thought it would be good to take a step back and get some vocabulary in place...
It's tempting to oversimplify things. Like neurons. It would be nice if there were one type of neuron, and all you needed to know about how neurons work could be clearly labelled on a diagram of that one type of neuron. Well, nature LOVES to specialize. So, before getting deeper into how neurons work, I thought it would be good to take a step back and get some vocabulary in place...
The Basics
From University of Washington's 'Neuroscience for kids': Neurons come in many different shapes and sizes. Some of the smallest neurons have cell bodies that are only 4 microns wide. Some of the biggest neurons have cell bodies that are 100 microns wide.
Neurons are similar to other cells in the body because:
- Neurons are surrounded by a cell membrane.
- Neurons have a nucleus that contains genes.
- Neurons contain cytoplasm, mitochondria and other "organelles".
- Neurons carry out basic cellular processes such as protein synthesis and energy production.
However, neurons differ from other cells in the body because:
- Neurons have specialized extensions called dendrites and axons. Dendrites bring information to the cell body and axons take information away from the cell body.
- Neurons communicate with each other through an electrochemical process.
- Neurons contain some specialized structures (for example, synapses) and chemicals (for example, neurotransmitters).
 From "Spiking Neuron Models" by Wulfram Gerstner and Werner M. Kistler (Cambridge University Press 2002): A typical neuron can be divided into three functionally distinct parts, called dendrites, soma, and axon. (see figure at right). Roughly speaking, the dendrites play the role of the `input device' that collects signals from other neurons and transmits them to the soma. The soma is the `central processing unit' that performs an important non-linear processing step: If the total input exceeds a certain threshold, then an output signal is generated. The output signal is taken over by the `output device', the axon, which delivers the signal to other neurons.
From "Spiking Neuron Models" by Wulfram Gerstner and Werner M. Kistler (Cambridge University Press 2002): A typical neuron can be divided into three functionally distinct parts, called dendrites, soma, and axon. (see figure at right). Roughly speaking, the dendrites play the role of the `input device' that collects signals from other neurons and transmits them to the soma. The soma is the `central processing unit' that performs an important non-linear processing step: If the total input exceeds a certain threshold, then an output signal is generated. The output signal is taken over by the `output device', the axon, which delivers the signal to other neurons.
It is common to refer to the sending neuron as the presynaptic cell and to the receiving neuron as the postsynaptic cell. A single neuron in vertebrate cortex often connects to more than 10,000 postsynaptic neurons. Many of its axonal branches end in the direct neighborhood of the neuron, but the axon can also stretch over several centimeters so as to reach to neurons in other areas of the brain.
Classes of Neurons
From University of Washington: One way to classify neurons is by the number of extensions that extend from the neuron's cell body (soma). 
Bipolar neurons have two processes extending from the cell body (examples: retinal cells, olfactory epithelium cells).
Pseudounipolar cells (example: dorsal root ganglion cells). Actually, these cells have 2 axons rather than an axon and dendrite. One axon extends centrally toward the spinal cord, the other axon extends toward the skin or muscle.
Multipolar neurons have many processes that extend from the cell body. However, each neuron has only one axon (examples: spinal motor neurons, pyramidal neurons, Purkinje cells).
Neurons can also be classified by the direction that they send information.
- Sensory (or afferent) neurons: send information from sensory receptors (e.g., in skin, eyes, nose, tongue, ears) TOWARD the central nervous system.
- Motor (or efferent) neurons: send information AWAY from the central nervous system to muscles or glands.
- Interneurons: send information between sensory neurons and motor neurons. Most interneurons are located in the central nervous system.
 From Wikipedia: Purkinje cells (or Purkinje neurons) are a class of GABAergic neuron located in the cerebellar cortex. They are named after their discoverer, Czech anatomist Jan Evangelista Purkyně.
From Wikipedia: Purkinje cells (or Purkinje neurons) are a class of GABAergic neuron located in the cerebellar cortex. They are named after their discoverer, Czech anatomist Jan Evangelista Purkyně.
These cells are some of the largest neurons in the human brain, with an intricately elaborate dendritic arbor, characterized by a large number of dendritic spines. Purkinje cells are found within the Purkinje layer in the cerebellum. Purkinje cells are aligned like dominos stacked one in front of the other. Their large dendritic arbors form nearly two dimensional layers through which parallel fibers from the deeper-layer granule cells pass. These parallel fibers make relatively weaker excitatory (glutamatergic) synapses to spines in the Purkinje cell dendrite, whereas climbing fibers originating from the inferior olivary nucleus in the medulla provide very powerful excitatory input to the proximal dendrites and cell soma. Parallel fibers pass orthogonally through the Purkinje neuron's dendritic arbor, with up to 200,000 parallel fibers forming a synapse with a single Purkinje cell. Alternatively, each Purkinje cell only receives a synapse from a single climbing fiber. Both basket and stellate cells (found in the cerebellar molecular layer) provide inhibitory (GABAergic) input to the Purkinje cell, with basket cells synapsing on the Purkinje cell axon initial segment and stellate cells onto the dendrites.
Purkinje cells send inhibitory projections to the deep cerebellar nuclei, and constitute the sole output of all motor coordination in the cerebellar cortex.
Purkinje cells show two distinct forms of electrophysiological activity:
- Simple spikes occur at rates of 50 - 150 Hz either spontaneously or and when Purkinje cells are activated synaptically by the parallel fibers, the axons of the granule cells.
- Complex spikes are rapid (>300 Hz) bursts of spikes caused by climbing fiber activation, and can involve the generation of calcium-mediated action potentials in the dendrites. Following complex spike activity simple spikes can be suppressed by the powerful complex spike input.
Purkinje cells show spontaneous electrophysiological activity in the form of trains of spikes, which may be important for cerebellar function.
Granule cells refer to tiny neurons (a type of cell) that are around 10 micrometres in diameter. Granule cells are found within the granular layer of the cerebellum, layer 4 of cerebral cortex, the dentate gyrus of the hippocampus, and in the olfactory bulb.
While anatomically similar, granule cells in different brain regions are functionally diverse. For instance, olfactory bulb granule cells are GABAergic and axonless, while granule cells in the dentate gyrus have glutamatergic projection axons. Interestingly, these two populations of granule cells are also the only major neuronal populations that undergo adult neurogenesis, while cerebellar and cortical granule cells do not.
Cerebellar granule cells account for nearly half of the neurons in the central nervous system. Granule cells receive excitatory input from mossy fibers originating from pontine nuclei. Cerebellar granule cells send parallel fibers up through the Purkinje layer into the molecular layer where they branch out and spread through Purkinje cell dendritic arbors. These parallel fibers form thousands of excitatory synapses with Purkinje cells.
Layer 4 granule cells of the cerebral cortex receive driving inputs from thalamus and convey driving inputs largely to supragranular layers 2-3, but also to infragranular layers of the cerebral cortex.
Golgi cells are inhibitory interneurons found within the granular layer of the cerebellum. These cells synapse onto the soma of granule cells. They receive excitatory input from mossy fibres, also synapsing on granule cells, and parallel fibers, which are long granule cell axons. Thereby this circuitry allows for feed-forward and feed-back inhibition of granule cells.
Wrapping things up
ref. Glial cells are nerve cells that don't carry nerve impulses. The various glial (meaning "glue") cells perform many important functions, including: digestion of parts of dead neurons, manufacturing myelin for neurons, providing physical and nutritional support for neurons, and more. Types of glial cells include Schwann's Cells, Satellite Cells, Microglia, Oligodendroglia, and Astroglia. Neuroglia (meaning "nerve glue") are the another type of brain cell. These cells guide neurons during fetal development.
Glial cells are not passive bystanders in cognition, however - they are active players. From "New Insights into Neuron-Glia Communication" by R. Douglas Fields and Beth Stevens-Graham (2002): Two-way communication between neurons and non-neural cells called glia is essential for axonal conduction, synaptic transmission, and information processing and thus is required for normal functioning of the nervous system during development and throughout adult life. The signals between neurons and glia include ion fluxes, neurotransmitters, cell adhesion molecules, and specialized signaling molecules released from synaptic and nonsynaptic regions of the neuron. In contrast to the serial flow of information along chains of neurons, glia communicate with other glial cells through intracellular waves of calcium and via intercellular diffusion of chemical messengers. By releasing neurotransmitters and other extracellular signaling molecules, glia can affect neuronal excitability and synaptic transmission and perhaps coordinate activity across networks of neurons.
 From Wikipedia: Schwann cells are a variety of neuroglia that mainly provide myelin insulation to axons in the peripheral nervous system of jawed vertebrates. The vertebrate nervous system relies on this myelin sheath for insulation and as a method of decreasing membrane capacitance in the axon, thus allowing for saltatory conduction to occur and for an increase in impulse speed, without an increase in axonal diameter. Non-myelinating Schwann cells are involved in maintenance of axons and are crucial for neuronal survival.
From Wikipedia: Schwann cells are a variety of neuroglia that mainly provide myelin insulation to axons in the peripheral nervous system of jawed vertebrates. The vertebrate nervous system relies on this myelin sheath for insulation and as a method of decreasing membrane capacitance in the axon, thus allowing for saltatory conduction to occur and for an increase in impulse speed, without an increase in axonal diameter. Non-myelinating Schwann cells are involved in maintenance of axons and are crucial for neuronal survival.
Schwann cells begin to form the myelin sheath in mammals during fetal development and work by spiraling around the axon, sometimes with as many as 100 revolutions. A well-developed Schwann cell is shaped like a rolled-up sheet of paper, with layers of myelin in between each coil. The inner layers of the wrapping, which are predominantly membrane material, form the myelin sheath while the outermost layer of nucleated cytoplasm forms the neurolemma. Only a small volume of residual cytoplasm communicates the inner from the outer layers. This is seen histologically as the Schmidt-Lantermann Incisure. Since each Schwann cell can cover about a millimeter (0.04 inches) along the axon, hundreds and often thousands are needed to completely cover an axon, which can sometimes span the length of a body. The gaps between the Schwann cell covered segments are the Nodes of Ranvier, important sites of ionic and other exchanges of the axon with the extracellular liquid. Unlike oligodendrocytes, myelinating Schwann cells provide insulation to only one axon (see image). This arrangement permits saltatory conduction of action potentials which greatly speeds it and saves energy.
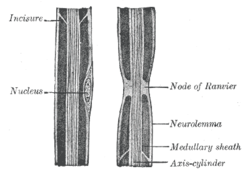 From Wikipedia: Nodes of Ranvier are regularly spaced gaps in the myelin sheath around an axon or nerve fiber. About one micrometer in length, these gaps expose the axonal membrane to the extracellular fluid. (The myelin sheath is the fatty tissue layer coating the axon.)
From Wikipedia: Nodes of Ranvier are regularly spaced gaps in the myelin sheath around an axon or nerve fiber. About one micrometer in length, these gaps expose the axonal membrane to the extracellular fluid. (The myelin sheath is the fatty tissue layer coating the axon.)
The myelin sheath helps speed the neural impulse by insulating the electrical current and making it possible for the impulse to jump from node to node, a process known as saltatory conduction, as opposed to traveling down the axon in tiny increments.
An action potential is the sharp electrochemical response of a stimulated neuron, a neuron whose membrane potential has been changed by a nearby cell, cells, or an experimentor. In an action potential, the cell membrane potential changes drastically and quickly as ions flow in or out of the cell. The action potential "travels" from one place in the cell to another, but ion flow occurs only at the nodes of Ranvier. Therefore, the action potential signal "jumps" along the axon, from node to node, rather than propagating smoothly, as they do in axons that lack a myelin sheath. This is due to clustering of voltage-gated Na+ and K+ ion channels at the Nodes of Ranvier.
Unmyelinated axons do not have Nodes of Ranvier; voltage gated ion channels in these axons are considerably less ordered and spread over the entire membrane surface.
Yale's Neuron Database
From Yale's Neuron Database for Vertebrates:
The Neuron Doctrine
The neuron doctrine is the now fundamental idea that neurons are the basic structural and functional units of the nervous system. The theory was put forward by Santiago Ramón y Cajal in the late 19th century. It held that neurons are discrete cells (not connected in a meshwork); that neurons are genetically and metabolically distinct units; that they have cell bodies, axons and dendrites; and that neural transmission goes only in one direction, from dendrites toward axons. (ref.)
The Neuron Doctrine, Redux
In the article "The Neuron Doctrine, Redux" (Bullock, T.H., Bennett, M.V.L., Johnston, D., Josephson, R., Marder, E., Fields R.D. 2005), the authors present a compelling case for expanding the scope of the original neuron doctrine.
 It turns out that electrical synapses are more common in the central nervous system than previously thought. They occur where the synaptic gap between two neurons is narrower than usual (2-4 nm), allowing special ion channels called 'connexons' that exist in the axon's synaptic region and the dendrite's synaptic region to connect to each other, creating ion channels that span the membrane of both the axon and dendrite. The ion channels allow the flow of polarizing ions and signaling molecules to flow directly from the transmitting neuron to the recieving neuron. (ref.)
It turns out that electrical synapses are more common in the central nervous system than previously thought. They occur where the synaptic gap between two neurons is narrower than usual (2-4 nm), allowing special ion channels called 'connexons' that exist in the axon's synaptic region and the dendrite's synaptic region to connect to each other, creating ion channels that span the membrane of both the axon and dendrite. The ion channels allow the flow of polarizing ions and signaling molecules to flow directly from the transmitting neuron to the recieving neuron. (ref.)
It also turns out that dendrites, like axons, have voltage-gated ion channels and can generate electrical potentials that carry information to and from the soma. This challenges the view that dendrites are simply passive recipients of information and axons the sole transmitters. It also suggests that the neuron is not simply active as a single element, but that complex computations can occur within a single neuron. (ref.)
And it's also important not to get too focused on single neurons. The single neuron spike record as indicator for a binary neural code persisted in mainstream neurophysiology until finally, in the late 1980s, it was challenged by the discovery of synchronized electrical wave electrical activity as a highly specific coordinating link among distributed neurons. (ref.)
Slow electrical potentials, action potentials initiated in dendrites, neuromodulatory effects, extrasynaptic release of neurotransmitters, and information flow between neurons and glia all contribute to information processing. (Science Magazine)
Summary
Ref: `Real' neurons are extremely complex biophysical and biochemical entities. Before designing a model it is therefore necessary to develop an intuition for what is important and what can be safely neglected. The Hodgkin-Huxley model describes the generation of action potentials on the level of ion channels and ion current flow. It is the starting point for detailed neuron models which in general include more than the three types of currents considered by Hodgkin and Huxley.
Electrophysiologists have described an overwhelming richness of different ion channels. The set of ion channels is different from one neuron to the next. The precise channel configuration in each individual neuron determines a good deal of its overall electrical properties. Synapses are usually modeled as specific ion channels that open for a certain time after presynaptic spike arrival.
The geometry of the neuron can play an important role in synaptic integration because the effect of synaptic input on the somatic membrane potential depends on the location of the synapses on the dendritic tree. Though some analytic results can be obtained for passive dendrites, it is usually necessary to resort to numerical methods and multi-compartment models in order to account for complex geometry and active ion channels.
Comments