 Overview
Overview
From Sandra Ackerman's book Discovering the Brain: The actual signals transmitted throughout the brain come in two forms, electrical and chemical. The two forms are interdependent and meet at the synapse, where chemical substances can alter the electrical conditions within and outside the cell membrane.
A nerve cell at rest holds a slight negative charge (about –70 millivolts, or thousandths of a volt, mV) with respect to the exterior; the cell membrane is said to be polarized. The negative charge, the resting potential of the membrane, arises from a very slight excess of negatively charged molecules inside the cell.
A membrane at rest is more or less impermeable to positively charged sodium ions (Na+), but when stimulated it is transiently open to their passage. The Na+ ions thus flow in, attracted by the negative charge inside, and the membrane temporarily reverses its polarity, with a higher positive charge inside than out. This stage lasts less than a millisecond, and then the sodium channels close again. Potassium channels (K+) open, and K+ ions move out through the membrane, reversing the flow of positively charged ions. (Both these channels are known as voltage-gated, meaning that they open or close in response to changes in electrical charge occurring across the membrane.)
Over the next 3 milliseconds, the membrane becomes slightly hyperpolarized, with a charge of about –80 mV, and then returns to its resting potential. During this time the sodium channels remain closed; the membrane is in a refractory phase.
An action potential—the very brief pulse of positive membrane voltage—is transmitted forward along the axon; it is prevented from propagating backward as long as the sodium channels remain closed. After the membrane has returned to its resting potential, however, a new impulse may arrive to evoke an action potential, and the cycle can begin again.
Gated channels, and the concomitant movement of ions in and out of the cell membrane, are widespread throughout the nervous system, with sodium, potassium, and chlorine being the most common ions involved. Calcium channels are also important, particularly at the presynaptic boutons of axons. When the membrane is at its resting potential, positively charged calcium ions (Ca2+) outside the cell far outnumber those inside. With the advent of an action potential, however, calcium ions rush into the cell. The influx of calcium ions leads to the release of neurotransmitter into the synaptic cleft; this passes the signal to a neighboring nerve cell.
Having taken a close look at the electrical side of the picture, we are in a better position to see where the chemistry comes in. Molecules of neurotransmitter are released into a synaptic cleft and bind to specific receptor sites on the postsynaptic side (the dendrite or dendritic spine), thereby altering the ion channels in the postsynaptic membrane. Some neurotransmitters cause sodium channels to open, allowing the influx of Na+ ions and thus a lessening of negative charge inside the cell membrane. If a considerable number of these potentials are received within a short interval, they can depolarize the membrane enough to trigger an action potential; the result is the transmission of a nerve impulse. The substances that can cause this to occur are the excitatory neurotransmitters. By contrast, other chemical compounds cause potassium channels to open, increasing the outflow of K+ ions from the cell and making excitation less likely; the neurotransmitters that bring about this state are considered inhibitory.
A given neuron has a great quantity of sites available on its dendrites and cell body and receives signals from many synapses simultaneously, both excitatory and inhibitory. These signals often amount to a rough balance; it is only when the net potential of the membrane in one region shifts significantly up or down from the resting level that a particular neurotransmitter can be said to be exerting an effect. Interestingly, in the membrane's overall balance sheet, the importance of a particular synapse varies with its proximity to where the axon leaves the nerve cell body, so that numerous excitatory potentials out at the ends of the dendrites may be overruled by several inhibitory potentials closer to the soma. Other kinds of synapse regulate the release of neurotransmitters into the synaptic cleft, where they go on to affect the postsynaptic channels as described above.
So, let's dig into this in more detail:
In every living cell there is always a difference in electrostatic potential between the inside and outside of a cell: the cell is polarized. (ref 1,ref 2). This membrane potential is due to ion pumps such as Na-K-ATP-ase, an enzyme. Within cells, the concentration of Na+ ions is lower, and that of K+ ions higher, than in the surrounding fluid. Na+, K+-ATPase and other ion pumps must work all the time in our body. If they were to stop, our cells would swell up, and might even burst, and we would rapidly lose consciousness. A great deal of energy is needed to drive ion pumps - in humans, about 1/3 of the ATP that the body produces.
The Na+, K+-ATPase enzyme exposes binding sites to 3 Na+ ions when opened to the inside of the cell, and binding sites to 2 K+ ions when opened to the outside of the cell, and uses the energy from ATP to change its shape from one configuration to the other. This results in a higher concentration of +ve ions outside of the cell than inside, as 3 Na+ ions are pumped out of the cell for every 2 K+ ions that are pumped into the cell. The resulting relative electric potential (voltage difference) is the driving 'electro motif force' (EMF) that causes ions to flow when ion channels, triggered by neurotransmitters, open.
From Fred Wolf's page at the Max Planck Institute for Dynamics and Self-Organization: Every living cell maintains a voltage difference across its cell membrane. (how this is done). Nerve cells distinguish themselves from other cells in that they use this voltage difference to process and transmit messages. When a nerve cell receives an impulse, the voltage across the cell membrane is reversed. This "action potential" spreads out through the long appendages of the cell with high speed. At the end of the appendages it is transmitted to other cells. In 1952, Alan Lloyd Hodgkin and Andrew Fielding Huxley described in a mathematical model how such an action potential originates on the basis of measurements on neurons of the squid. The Hodgkin-Huxley model, for which the scientists received the 1963 Nobel Prize in Medicine, has since then served to explain the signal processes in all neurons.
According to the Hodgkin-Huxley model, an action potential is initiated when the voltage across the membrane of the nerve cell reaches to a certain threshold value. Voltage gated sodium channels react to this voltage change by opening up and triggering an avalanche-like reaction. Positively charged sodium ions flow through the open channels into the cell, which leads to a further increase of the membrane potential and the opening of additional sodium channels. The threshold and the speed with which the action potential originates vary from cell to cell - for any individual cell however, these parameters are specified for the most part by the characteristics of its sodium channels.
There is always a difference in electrostatic potential between the inside and outside of a cell: the cell is polarized. the result of the distribution of ions across the cell membrane and the permeability of the membrane to these ions. The ion channels themselves are positioned and held in place by the cytoskeleton of the neuron. In fact the Baz/Par-6/aPKC complex, a highly conserved protein cassette that functions during the establishment of polarity in a number of cell types, is also involved in the development of new synaptic boutons. (ref.)
From the University of Bristol: How do neurons work? : Under resting conditions, the potassium channel is more permeable to potassium ions than the sodium channel is to sodium ions. So there is a slow outward leak of potassium ions that is larger than the inward leak of sodium ions. This means that the membrane has a charge on the inside face that is negative relative to the outside, as more positively charged ions flow out of the neuron than flow in. The membrane is said to be polarised.
From reference.com: The voltage of an inactive cell remains close to a resting potential with excess negative charge inside the cell. When the membrane of an excitable cell becomes depolarized beyond a threshold, the cell undergoes an action potential (it "fires), often called a "spike" (see Threshold and initiation).
An action potential is a rapid change of the polarity of the voltage from negative to positive and then vice versa, the entire cycle lasting on the order of milliseconds. Each cycle — and therefore each action potential — has a rising phase, a falling phase, and finally an undershoot (see Phases).
From http://www.du.edu/~kinnamon/3640/actionpotential/ap1.html : Conformational changes in the voltage-dependent Na+ channel constitute the basis of the action potential. Time course of an action potential: The opening of a few Na+ channels leads to an initial depolarization. If this depolarization exceeds a threshold value then a rapid regenerative opening of many other Na+ channels follows, resulting in the depolarizing phase of an action potential. Since the Na+ channel spontaneously inactivates, the repolarization of the membrane occurs automatically. (Reichert, Introduction to Neurobiology)
From the commentary by Frank Werblin, UC Berkely to his animated and interactive tutorial's action potential animation at http://mcb.berkeley.edu/courses/mcb64/action_potential.html :
 The action potential begins with a small depoloarization that comes from somewhere else. This causes ALL ion gates involved to begin their openning trajectory, but each at a different rate. First the sodium activation gate at the top of the sodium channel begins to open, allowing the sodium to enter the membrane and depolarize it further. Then the sodium inactivation gate at the bottom of the sodium channel closes, terminating the inward flow of sodium.
The action potential begins with a small depoloarization that comes from somewhere else. This causes ALL ion gates involved to begin their openning trajectory, but each at a different rate. First the sodium activation gate at the top of the sodium channel begins to open, allowing the sodium to enter the membrane and depolarize it further. Then the sodium inactivation gate at the bottom of the sodium channel closes, terminating the inward flow of sodium.
More slowly, the potassium activation gate begins to open. There's an outward flow of potassium ions, hyperpolarizing the membrane. This outward flow continues until the membrane overshoots the resting level in the hyperpolarizing direction. The potassium gate opens with depolarization, but the current hyperpolarization causes the potassium gate to close, terminating the action potential.
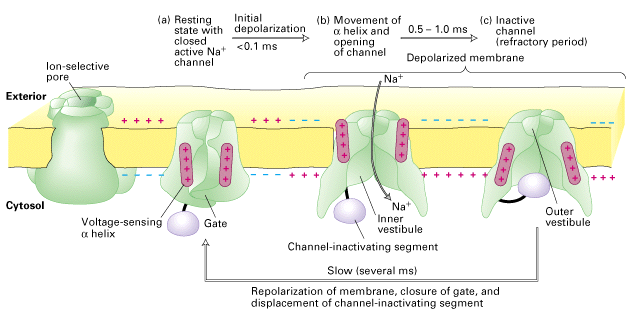
The picture at right (from Figure 21-13 Molecular Cell Biology, 4th edition OR Figure 7-33 Molecular Cell Biology, 5th edition, by Lodish et al) shows the structure and function of the voltage-gated Na+ channel. More detailed information on this process is provided by http://www.zoology.ubc.ca/~gardner/action_potential.htm
Bucket Brigade
 (The diagrams at right are from B. Fleming, U. of Waterloo )
(The diagrams at right are from B. Fleming, U. of Waterloo )
From the University of Bristol: How do neurons work? :
Excess ions are subsequently pumped in/out of the neuron.
In some neurons, after hyperpolarization a secondary, longer-term effect occurs: a post-excitatory restoration of membrane potential, termed after-hyperpolarization (AHP).
If this were all there was to it, then the action potential would propagate in all directions along an axon. But action potentials move in one direction. This is achieved because the sodium channels have a refactory period following activation, during which they cannot open again. This ensures that the action potential is proagated in a specific direction along the axon.
For a neuron to integrate all of the various pulses that it is recieving from various other neurons, the pulses need to 'line up' to a certain degree in order to be 'coincident' and push the neuron over the activation threshold. For this to happen, the pulse width would need to be greater than the propagation delay introduced by the axon. The pulse width is on the order of 2ms. The propagation along an unmyelinated axon depends on a number of factors (e.g. the cross-sectional width of the axon, the type of axon, etc.) but is typically around 50 m/s. (ref.) which would imply that the axon needs to be less than 10 cm in length.
 Axons that are used a lot become sheathed in 1 mm long sections of myelin (oligodendrocytes) that promote more efficient and faster conduction. The action potential in myelinated axons jumps from one gap in the myelin (called a 'node of Ranvier') to the next in a process called 'saltatory conduction'. Myelination causes the resistance of the membrane to increase by a factor of 200 and the capacitance per unit area of the axon to decrease by a factor of 200 (ref.)
Axons that are used a lot become sheathed in 1 mm long sections of myelin (oligodendrocytes) that promote more efficient and faster conduction. The action potential in myelinated axons jumps from one gap in the myelin (called a 'node of Ranvier') to the next in a process called 'saltatory conduction'. Myelination causes the resistance of the membrane to increase by a factor of 200 and the capacitance per unit area of the axon to decrease by a factor of 200 (ref.)
Recent research by B. Stevens, S. Porta, L. Haak, V. Gallo and R. Douglas Fields has shown that axons release Adenosine in response to action potential firing and that this is used as a chemical signal by oligodendrocyte progenitor cells to initiate sheathing axons that fire frequently.
More information:
MASS ACTION IN THE NERVOUS SYSTEM EXAMINATION OF THE NEUROPHYSIOLOGICAL BASIS OF ADAPTIVE BEHAVIOR THROUGH THE EEG (2004) by Professor Walter J. Freeman University of California, Berkeley, USA (http://sulcus.berkeley.edu/MANSWWW/MANSHTML/MANSChapt3.html) : provides a lot of the equations and details about how neurons operate.
Comments